Структура одного белка определяется группой. Белки структура белковой молекулы l l выяснение. Локальное упорядочивание участка цепи
Доказано существование 4 уровней структурной организации белковой молекулы.
Первичная структура белка – последовательность расположения аминокислотных остатков в полипептидной цепи. В белках отдельные аминокислоты связаны друг с другом пептидными связями , возникающими при взаимодействии a-карбоксильных и a-аминогрупп аминокислот.
К настоящему времени расшифрована первичная структура десятков тысяч разных белков. Для определения первичной структуры белка методами гидролиза выясняют аминокислотный состав. Затем определяют химическую природу концевых аминокислот. Следующий этап - определение последовательности аминокислот в полипептидной цепи. Для этого используют избирательный частичный (химический и ферментативный) гидролиз. Возможно применение рентгеноструктурного анализа, а также данных о комплементарной нуклеотидной последовательности ДНК.
Вторичная структура белка – конфигурация полипептидной цепи, т.е. способ упаковки полипептидной цепи в определенную конформацию. Процесс этот протекает не хаотично, а в соответствии с программой, заложенной в первичной структуре.
Стабильность вторичной структуры обеспечивается в основном водородными связями, однако определенный вклад вносят ковалентные связи – пептидные и дисульфидные.
Наиболее вероятным типом строения глобулярных белков считают a-спираль . Закручивание полипептидной цепи происходит по часовой стрелке. Для каждого белка характерна определенная степень спирализации. Если цепи гемоглобина спирализованы на 75%, то пепсина-всего на 30%.
Тип конфигурации полипептидных цепей, обнаруженных в белках волос, шелка, мышц, получил название b-структуры . Сегменты пептидной цепи располагаются в один слой, образуя фигуру, подобную листу, сложенному в гармошку. Слой может быть образован двумя или большим количеством пептидных цепей.
В природе существуют белки, строение которых не соответствует ни β-, ни a-структуре, например, коллаген - фибриллярный белок, составляющий основную массу соединительной ткани в организме человека и животных.
Третичная структура белка – пространственная ориентация полипептидной спирали или способ укладки полипептидной цепи в определенном объеме. Первый белок, третичная структура которого была выяснена рентгеноструктурным анализом - миоглобин кашалота (рис. 2).
В стабилизации пространственной структуры белков, помимо ковалентных связей, основную роль играют нековалентные связи (водородные, электростатические взаимодействия заряженных групп, межмолекулярные ван-дер-ваальсовы силы, гидрофобные взаимодействия и т.д.).
По современным представлениям, третичная структура белка после завершения его синтеза формируется самопроизвольно. Основной движущей силой является взаимодействие радикалов аминокислот с молекулами воды. При этом неполярные гидрофобные радикалы аминокислот погружаются внутрь белковой молекулы, а полярные радикалы ориентируются в сторону воды. Процесс формирование нативной пространственной структуры полипептидной цепи называют фолдингом . Из клеток выделены белки, названные шаперонами. Они участвуют в фолдинге. Описан ряд наследственных заболеваний человека, развитие которых связывают с нарушением вследствие мутаций процесса фолдинга (пигментозы, фиброзы и др.).
Методами рентгеноструктурного анализа доказано существование уровней структурной организации белковой молекулы, промежуточных между вторичной и третичной структурами. Домен - это компактная глобулярная структурная единица внутри полипептидной цепи (рис. 3). Открыто много белков (например, иммуноглобулины), состоящих из разных по структуре и функциям доменов, кодируемых разными генами.
Все биологические свойства белков связаны с сохранностью их третичной структуры, которую называют нативной . Белковая глобула не является абсолютно жесткой структурой: возможны обратимые перемещения частей пептидной цепи. Эти изменения не нарушают общей конформации молекулы. На конформацию молекулы белка оказывают влияние рН среды, ионная сила раствора, взаимодействие с другими веществами. Любые воздействия, приводящие к нарушению нативной конформации молекулы, сопровождаются частичной или полной потерей белком его биологических свойств.
Четвертичная структура белка - способ укладки в пространстве отдельных полипептидных цепей, обладающих одинаковой или разной первичной, вторичной или третичной структурой, и формирование единого в структурном и функциональном отношениях макромолекулярного образования.
Белковую молекулу, состоящую из нескольких полипептидных цепей, называют олигомером , а каждую входящую в него цепь - протомером . Олигомерные белки чаще построены из четного числа протомеров, например, молекула гемоглобина состоит из двух a- и двух b-полипептидных цепей (рис. 4).
Четвертичной структурой обладает около 5% белков, в том числе гемоглобин, иммуноглобулины. Субъединичное строение свойственно многим ферментам.
Белковые молекулы, входящие в состав белка с четвертичной структурой, образуются на рибосомах по отдельности и лишь после окончания синтеза образуют общую надмолекулярную структуру. Биологическую активность белок приобретает только при объединении входящих в его состав протомеров. В стабилизации четвертичной структуры принимают участие те же типы взаимодействий, что и в стабилизации третичной.
Некоторые исследователи признают существование пятого уровня структурной организации белков. Это метаболоны - полифункциональные макромолекулярные комплексы разных ферментов, катализирующих весь путь превращений субстрата (синтетазы высших жирных кислот, пируватдегидрогеназный комплекс, дыхательная цепь).
Первичной структурой белков называется линейная полипептидная цепь из аминокислот, соединенных между собой пептидными связями. Первичная структура - простейший уровень структурной организации белковой молекулы. Высокую стабильность ей придают ковалентные пептидные связи между α-аминогруппой одной аминокислоты и α-карбоксильной группой другой аминокислоты
Если в образовании пептидной связи участвует иминогруппа пролина или гидроксипролина, то она имеет другой вид

При образовании пептидных связей в клетках сначала активируется карбоксильная группа одной аминокислоты, а затем она соединяется с аминогруппой другой. Примерно так же проводят лабораторный синтез полипептидов.
Пептидная связь является повторяющимся фрагментом полипептидной цепи. Она имеет ряд особенностей, которые влияют не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи:


· копланарность - все атомы, входящие в пептидную группу, находятся в одной плоскости;
· способность существовать в двух резонансных формах (кето- или енольной форме);
· транс-положение заместителей по отношению к С-N-связи;
· способность к образованию водородных связей, причем каждая из пептидных групп может образовывать две водородные связи с другими группами, в том числе и пептидными.
Исключение составляют пептидные группы с участием аминогруппы пролина или гидроксипролина. Они способны образовывать только одну водородную связь (см. выше). Это сказывается на формировании вторичной структуры белка. Полипептидная цепь на участке, где находится пролин или гидроксипролин, легко изгибается, так как не удерживается, как обычно, второй водородной связью.
схема образования трипептида:

Уровни пространственной организации белков: вторичная структура белков: понятие об α-спирали и β-складчатом слое. Третичная структура белков: понятие о нативном белке и денатурации белка. Четвертичная структура белков на примере строения гемоглобина.
Вторичная структура белка. Под вторичной структурой белка понимают способ укладки полипептидной цепи в упорядоченную структуру. По конфигурации выделяют следующие элементы вторичной структуры: α -спираль и β -складчатый слой.
Модель строения α-спирали, учитывающая все свойства пептидной связи, была разработана Л. Полингом и Р. Кори (1949 - 1951 гг.).
На рисунке 3, а изображена схема α -спирали, дающая представление об основных ее параметрах. Полипептидная цепь сворачивается вα -спираль таким образом, что витки спирали регулярны, поэтому спиральная конфигурация имеет винтовую симметрию (рис. 3, б ). На каждый виток α -спирали приходится 3,6 аминокислотных остатка. Расстояние между витками или шаг спирали составляет 0,54 нм, угол подъема витка равен 26°. Формирование и поддержание α -спиральной конфигурации происходит за счет водородных связей, образующихся между пептидными группами каждого n -го и (п + 3)-го аминокислотных остатков. Хотя энергия водородных связей мала, большое количество их приводит к значительному энергетическому эффекту, в результате чего α -спиральная конфигурация довольно устойчива. Боковые радикалы аминокислотных остатков не участвуют в поддержании α -спиральной конфигурации, поэтому все аминокислотные остатки в α -спирали равнозначны.
В природных белках существуют только правозакрученные α -спирали.
β-Складчатый слой - второй элемент вторичной структуры. В отличие от α -спирали β -складчатый слой имеет линейную, а не стержневую форму (рис. 4). Линейная структура удерживается благодаря возникновению водородных связей между пептидными группировками, стоящими на разных участках полипептидной цепи. Эти участки оказываются сближенными на расстояние водородной связи между - С = О и HN - группами (0,272 нм).

Рис. 4. Схематичное изображение β
-складчатого слоя (стрелками указан
 о направление полипептидной цепи)
о направление полипептидной цепи)
Рис. 3. Схема (а ) и модель (б ) α -спирали
Вторичная структура белка определяется первичной. Аминокислотные остатки в разной степени способны к образованию водородных связей, это и влияет на образование α -спирали или β -слоя. К спиралеобразующим аминокислотам относятся аланин, глутаминовая кислота, глутамин, лейцин, лизин, метионин и гистидин. Если фрагмент белка состоит главным образом из перечисленных выше аминокислотных остатков, то на данном участке сформируется α -спираль. Валин, изолейцин, треонин, тирозин и фенилаланин способствуют образованию β -слоев полипептидной цепи. Неупорядоченные структуры возникают на участках полипептидной цепи, где сконцентрированы такие аминокислотные остатки, как глицин, серии, аспарагиновая кислота, аспарагин, пролин.
Во многих белках одновременно имеются и α -спирали, и β -слои. Доля спиральной конфигурации у разных белков различна. Так, мышечный белок парамиозин практически на 100% спирализован; высока доля спиральной конфигурации у миоглобина и гемоглобина (75%). Напротив, у трипсина и рибонуклеазы значительная часть полипептидной цепи укладывается в слоистые β -структуры. Белки опорных тканей - кератин (белок волос), коллаген (белок кожи и сухожилий) - имеют β -конфигурацию полипептидных цепей.
Третичная структура белка. Третичная структура белка - это способ укладки полипептидной цепи в пространстве. Чтобы белок приобрел присущие ему функциональные свойства, полипептидная цепь должна определенным образом свернуться в пространстве, сформировав функционально активную структуру. Такая структура называется нативной. Несмотря на громадное число теоретически возможных для отдельной полипептидной цепи пространственных структур, сворачивание белка приводит к образованию единственной нативной конфигурации.
Стабилизируют третичную структуру белка взаимодействия, возникающие между боковыми радикалами аминокислотных остатков разных участков полипептидной цепи. Эти взаимодействия можно разделить на сильные и слабые.
К сильным взаимодействиям относятся ковалентные связи между атомами серы остатков цистеина, стоящих в разных участках полипептидной цепи. Иначе такие связи называются дисульфидными мостами; образование дисульфидного моста можно изобразить следующим образом:
Кроме ковалентных связей третичная структура белковой молекулы поддерживается слабыми взаимодействиями, которые, в свою очередь, разделяются на полярные и неполярные.
К полярным взаимодействиям относятся ионные и водородные связи. Ионные взаимодействия образуются при контакте положительно заряженных групп боковых радикалов лизина, аргинина, гистидина и отрицательно заряженной СООН-группы аспарагиновой и глутаминовой кислот. Водородные связи возникают между функциональными группами боковых радикалов аминокислотных остатков.
Неполярные или ван-дер-ваальсовы взаимодействия между углеводородными радикалами аминокислотных остатков способствуют формированию гидрофобного ядра (жирной капли) внутри белковой глобулы, т.к. углеводородные радикалы стремятся избежать соприкосновения с водой. Чем больше в составе белка неполярных аминокислот, тем большую роль в формировании его третичной структуры играют ван-дер-ваальсовы связи.
Многочисленные связи между боковыми радикалами аминокислотных остатков определяют пространственную конфигурацию белковой молекулы (рис. 5).

Рис. 5. Типы связей, поддерживающих третичную структуру белка:
а
- дисульфидный мостик; б -
ионная связь; в, г -
водородные связи;
д -
ван-дер-ваальсовы связи
Третичная структура отдельно взятого белка уникальна, как уникальна и его первичная структура. Только правильная пространственная укладка белка делает его активным. Различные нарушения третичной структуры приводят к изменению свойств белка и потере биологической активности.
Четвертичная стурктура белка. Белки с молекулярной массой более 100 кДа 1 состоят, как правило, из нескольких полипептидных цепей со сравнительно небольшой молекулярной массой. Структура, состоящая из определенного числа полипептидных цепей, занимающих строго фиксированное положение относительно друг друга, вследствие чего белок обладает той или иной активностью, называется четвертичной структурой белка. Белок, обладающий четвертичной структурой, называется эпимолекулой илимультимером , а составляющие его полипептидные цепи - соответственно субъединицами или протомерами . Характерным свойством белков с четвертичной структурой является то, что отдельная субъединица не обладает биологической активностью.
Стабилизация четвертичной структуры белка происходит за счет полярных взаимодействий между боковыми радикалами аминокислотных остатков, локализованных на поверхности субъединиц. Такие взаимодействия прочно удерживают субъединицы в виде организованного комплекса. Участки субъединиц, на которых происходят взаимодействия, называются контактными площадками.
Классическим примером белка, имеющего четвертичную структуру, является гемоглобин. Молекула гемоглобина с молекулярной массой 68 000 Да состоит из четырех субъединиц двух разных типов - α и β / α -Субъединица состоит из 141 аминокислотного остатка, a β - из 146. Третичная стурктура α - и β -субъединиц сходна, как и их молекулярная масса (17 000 Да). Каждая субъединица содержит простетическую группу - гем . Поскольку гем присутствует и в других белках (цитохромы, миоглобин), которые будут изучаться далее, хотя бы коротко обсудим структуру тема (рис. 6). Группировка гема представляет собой сложную копланарную циклическую систему, состоящую из центрального атома, который образует координационные связи с четырьмя остатками пиррола, соединенными метановыми мостиками (= СН -). В гемоглобине железо обычно находится в состоянии окисления (2+).
Четыре субъединицы - две α и две β - соединяются в единую структуру таким образом, что α -субъединицы контактируют только с β -субъединицами и наоборот (рис. 7).

Рис. 6. Структура гема гемоглобина

Рис. 7. Схематичное изображение четвертичной структуры гемоглобина:
Fe - гем гемоглобина
Как видно из рисунка 7, одна молекула гемоглобина способна переносить 4 молекулы кислорода. И связывание, и освобождение кислорода сопровождается конформационными изменениями структуры α - и β -субъединиц гемоглобина и их взаимного расположения в эпимолекуле. Этот факт свидетельствует о том, что четвертичная структура белка не является абсолютно жесткой.
Похожая информация.
 l l Выяснение структурной организации белков – одна из главных проблем современной биохимии Оно имеет важное научно-практическое значение для понимания огромного разнообразия функций белков Белковые молекулы – продукт поликонденсации 20 -ти различных аминокислот, соединенных не хаотично, а в строгом соответствии с кодом белкового синтеза Каким образом соединяются между собой многие десятки и сотни аминокислот в белковой молекуле?
l l Выяснение структурной организации белков – одна из главных проблем современной биохимии Оно имеет важное научно-практическое значение для понимания огромного разнообразия функций белков Белковые молекулы – продукт поликонденсации 20 -ти различных аминокислот, соединенных не хаотично, а в строгом соответствии с кодом белкового синтеза Каким образом соединяются между собой многие десятки и сотни аминокислот в белковой молекуле?
 Полипептидная теория строения белка l l 1902 г. , Эмиль Фишер Белки – сложные полипептиды, в которых отдельные аминокислоты связаны друг с другом пептидными (R–CO–NH–R) связями, возникающими при взаимодействии карбоксильных и аминогрупп аминокислот
Полипептидная теория строения белка l l 1902 г. , Эмиль Фишер Белки – сложные полипептиды, в которых отдельные аминокислоты связаны друг с другом пептидными (R–CO–NH–R) связями, возникающими при взаимодействии карбоксильных и аминогрупп аминокислот
 Экспериментальные доказательства теории l l l В природных белках сравнительно мало свободных групп –СООН и –NH 2 В процессе гидролиза белка образуется стехиометрическое количество титруемых групп – СООН и –NH 2 Под действием протеолитических ферментов белки расщепляются на строго определенные фрагменты (полипептиды) с концевыми аминокислотами, соответствующими избирательности действия ферментов Белки дают биуретовую реакцию Анализ рентгенограмм подтверждает полипептидную структуру белка Существенное подтверждение полипептидной теории строения белка – возможность синтеза химическими методами полипептидов и белков с уже известным строением (инсулина, лизоцима, рибонуклеазы и др.)
Экспериментальные доказательства теории l l l В природных белках сравнительно мало свободных групп –СООН и –NH 2 В процессе гидролиза белка образуется стехиометрическое количество титруемых групп – СООН и –NH 2 Под действием протеолитических ферментов белки расщепляются на строго определенные фрагменты (полипептиды) с концевыми аминокислотами, соответствующими избирательности действия ферментов Белки дают биуретовую реакцию Анализ рентгенограмм подтверждает полипептидную структуру белка Существенное подтверждение полипептидной теории строения белка – возможность синтеза химическими методами полипептидов и белков с уже известным строением (инсулина, лизоцима, рибонуклеазы и др.)
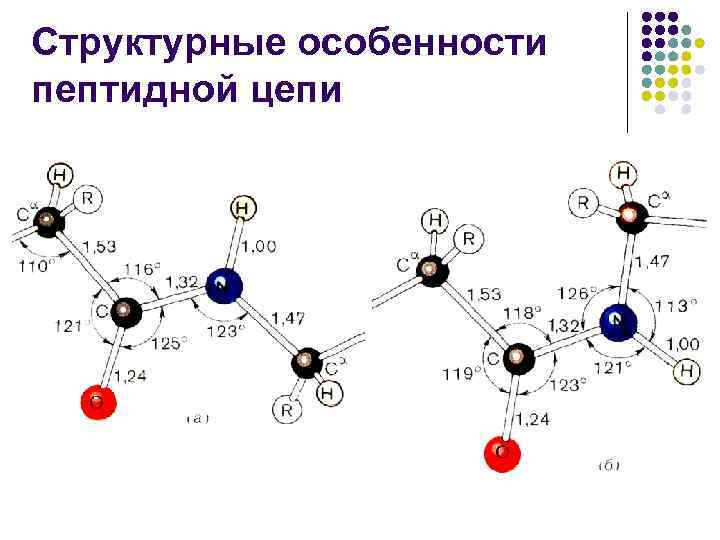
 Структурные особенности пептидной цепи l l Пептидная связь примерно на 10% короче связи –С–N– и имеет характер «частично двойной» связи –С=N– Л. Полинг и Р. Кори предложили в 1948– 1955 гг. объяснять особую природу связи С–N «резонансом» между двумя формами
Структурные особенности пептидной цепи l l Пептидная связь примерно на 10% короче связи –С–N– и имеет характер «частично двойной» связи –С=N– Л. Полинг и Р. Кори предложили в 1948– 1955 гг. объяснять особую природу связи С–N «резонансом» между двумя формами
 Структурные особенности пептидной цепи l l l Другими словами, в белках и пептидах связь С–N является частично кратной из-за взаимодействия неподеленной пары электронов атома азота с электронной системой карбонильной группы, что приводит к затрудненному вращению вокруг связи С–N Обычно пептидная связь имеет транс-конфигурацию, что выгоднее цис-конфигурации на 2, 6 ккал/моль (10, 878 к. Дж/моль), т. к. сближение -углеродных атомов в цисконфигурации вызывает пространственные затруднения Эта разница в значительной мере снимается, если азот пептидной связи принадлежит про
Структурные особенности пептидной цепи l l l Другими словами, в белках и пептидах связь С–N является частично кратной из-за взаимодействия неподеленной пары электронов атома азота с электронной системой карбонильной группы, что приводит к затрудненному вращению вокруг связи С–N Обычно пептидная связь имеет транс-конфигурацию, что выгоднее цис-конфигурации на 2, 6 ккал/моль (10, 878 к. Дж/моль), т. к. сближение -углеродных атомов в цисконфигурации вызывает пространственные затруднения Эта разница в значительной мере снимается, если азот пептидной связи принадлежит про
 Структурные особенности пептидной цепи l l l Повороты возможны вокруг двух простых связей (N–С и С –C 1), примыкающих к асимметрическому атому Такие повороты измеряются двугранными углами и Любые конформации пептидной цепи могут быть описаны набором значений углов и у каждого из С -атомов (обычно = 180 – транс-форма)
Структурные особенности пептидной цепи l l l Повороты возможны вокруг двух простых связей (N–С и С –C 1), примыкающих к асимметрическому атому Такие повороты измеряются двугранными углами и Любые конформации пептидной цепи могут быть описаны набором значений углов и у каждого из С -атомов (обычно = 180 – транс-форма)
 Структурные особенности пептидной цепи l l В силу взаимодействия между заместителями в пептидной цепи углы и не могут принимать любые значения – для них разрешенными оказываются лишь некоторые дискретные области, соответствующие энергетически выгодным конформациям пептидной цепи Углы и взаимосвязаны, изменение одного из них влечет изменение второго Гопалачамудрам Нарайана Рамачандран
Структурные особенности пептидной цепи l l В силу взаимодействия между заместителями в пептидной цепи углы и не могут принимать любые значения – для них разрешенными оказываются лишь некоторые дискретные области, соответствующие энергетически выгодным конформациям пептидной цепи Углы и взаимосвязаны, изменение одного из них влечет изменение второго Гопалачамудрам Нарайана Рамачандран

 l К. У. Линдерстрем-Ланг предложил различать 4 уровня организации белковых молекул – первичную, вторичную, третичную и четвертичную структуры. Хотя эти категории в известной степени устарели, ими пока продолжают пользоваться
l К. У. Линдерстрем-Ланг предложил различать 4 уровня организации белковых молекул – первичную, вторичную, третичную и четвертичную структуры. Хотя эти категории в известной степени устарели, ими пока продолжают пользоваться

 Первичная структура белка l l l последовательность аминокислотных остатков в полипептидной цепи кодируется структурным геном данного белка и содержит в себе все необходимое для самоорганизации его пространственной структуры Все белки различаются по своей первичной структуре Потенциально возможное число таких структур неограниченно Однако в живой природе все эти возможности не реализуются: общее число различных типов белков у всех видов живых организмов составляет величину порядка 1010– 1012
Первичная структура белка l l l последовательность аминокислотных остатков в полипептидной цепи кодируется структурным геном данного белка и содержит в себе все необходимое для самоорганизации его пространственной структуры Все белки различаются по своей первичной структуре Потенциально возможное число таких структур неограниченно Однако в живой природе все эти возможности не реализуются: общее число различных типов белков у всех видов живых организмов составляет величину порядка 1010– 1012
 l l l Познание биологической функции и, в частности, молекулярного механизма физиологического действия белка невозможно без детального знания его строения Установление первичной структуры белка – основа для определения вторичной и третичной структур, выяснения расположения функциональных групп в его активном центре, путь к познанию механизма его функционирования. Исследование первичной структуры «мутагенных» белков позволяет на молекулярном уровне выяснить характер наследственных болезней Данные по первичной структуре используются как один из показателей при установлении и проверке таксономических взаимоотношений между различными видами живых организмов и построении схемы биологической эволюции
l l l Познание биологической функции и, в частности, молекулярного механизма физиологического действия белка невозможно без детального знания его строения Установление первичной структуры белка – основа для определения вторичной и третичной структур, выяснения расположения функциональных групп в его активном центре, путь к познанию механизма его функционирования. Исследование первичной структуры «мутагенных» белков позволяет на молекулярном уровне выяснить характер наследственных болезней Данные по первичной структуре используются как один из показателей при установлении и проверке таксономических взаимоотношений между различными видами живых организмов и построении схемы биологической эволюции
 Методы определения первичной структуры l l Подготовка белка к анализу первичной структуры призвана свести к минимуму влияние других, более высоких уровней его организации Иными словами, объектом анализа должна быть неупорядоченная белковая цепь без каких-либо поперечных ковалентных связей (например, дисульфидных) так, чтобы все ее звенья, все пептидные связи были в равной мере доступны действию как химических реагентов, так и ферментов
Методы определения первичной структуры l l Подготовка белка к анализу первичной структуры призвана свести к минимуму влияние других, более высоких уровней его организации Иными словами, объектом анализа должна быть неупорядоченная белковая цепь без каких-либо поперечных ковалентных связей (например, дисульфидных) так, чтобы все ее звенья, все пептидные связи были в равной мере доступны действию как химических реагентов, так и ферментов
 l l Белок должен быть прежде всего подвергнут глубокой денатурации и утратить четвертичную, третичную и по возможности вторичную структуры Если в нем имеются дисульфидные связи, прибегают к их расщеплению, используя, как правило, восстановление большим избытком меркаптосоединения
l l Белок должен быть прежде всего подвергнут глубокой денатурации и утратить четвертичную, третичную и по возможности вторичную структуры Если в нем имеются дисульфидные связи, прибегают к их расщеплению, используя, как правило, восстановление большим избытком меркаптосоединения
 l Во избежание замыкания дисульфидных связей вновь сульфгидрильные группы блокируют избытком иодуксусной кислоты: R–SН + I–СН 2–СООН R–S–СН 2–СООН + НI
l Во избежание замыкания дисульфидных связей вновь сульфгидрильные группы блокируют избытком иодуксусной кислоты: R–SН + I–СН 2–СООН R–S–СН 2–СООН + НI
 Затем проводят селективный гидролиз денатурированного белка для получения системы перекрывающихся пептидов. Гидролиз проводят ферментами: l l l трипсином (связи, образованные карбоксильными группами арг или лиз), химотрипсином (связи, образованные группами –СООН ароматических аминокислот) и др. ; химическими реагентами: бромцианом Br –С N (связи, образованные группами –СООН мет) и др.
Затем проводят селективный гидролиз денатурированного белка для получения системы перекрывающихся пептидов. Гидролиз проводят ферментами: l l l трипсином (связи, образованные карбоксильными группами арг или лиз), химотрипсином (связи, образованные группами –СООН ароматических аминокислот) и др. ; химическими реагентами: бромцианом Br –С N (связи, образованные группами –СООН мет) и др.
 l l l Следующий этап – фракционирование полученных пептидов – осуществляется методами хроматографии Наконец, последний этап – определение первичной структуры l начинается с определения его молекулярной массы, аминокислотного состава, N- и Сконцевых аминокислотных остатков После того, как структура всех фрагментов установлена, белок подвергают расщеплению при помощи другого агента и получают второй, отличный от первого набор пептидных фрагментов, которые разделяют и анализируют для выяснения порядка расположения аминокислот в белке
l l l Следующий этап – фракционирование полученных пептидов – осуществляется методами хроматографии Наконец, последний этап – определение первичной структуры l начинается с определения его молекулярной массы, аминокислотного состава, N- и Сконцевых аминокислотных остатков После того, как структура всех фрагментов установлена, белок подвергают расщеплению при помощи другого агента и получают второй, отличный от первого набор пептидных фрагментов, которые разделяют и анализируют для выяснения порядка расположения аминокислот в белке

 Метод Ф. Сенджера l В 1945 г. английский биохимик Ф. Сенджер предложил один из первых методов определения N-концевых аминокислот
Метод Ф. Сенджера l В 1945 г. английский биохимик Ф. Сенджер предложил один из первых методов определения N-концевых аминокислот
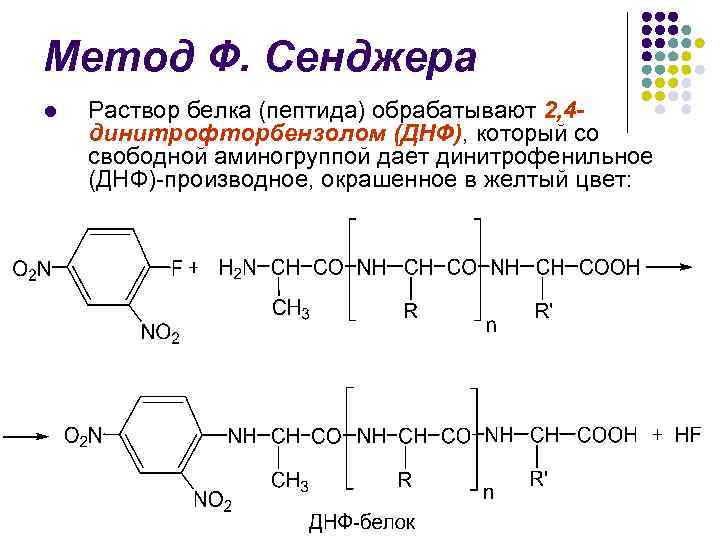 Метод Ф. Сенджера l Раствор белка (пептида) обрабатывают 2, 4 динитрофторбензолом (ДНФ), который со свободной аминогруппой дает динитрофенильное (ДНФ)-производное, окрашенное в желтый цвет:
Метод Ф. Сенджера l Раствор белка (пептида) обрабатывают 2, 4 динитрофторбензолом (ДНФ), который со свободной аминогруппой дает динитрофенильное (ДНФ)-производное, окрашенное в желтый цвет:
 Метод Ф. Сенджера l l Последующий кислотный гидролиз (5, 7 н. НСl) приводит к разрыву пептидных связей и образованию ДНФ-производного Nконцевой аминокислоты Последнее экстрагируется эфиром и идентифицируется методом ТСХ в присутствии стандартов
Метод Ф. Сенджера l l Последующий кислотный гидролиз (5, 7 н. НСl) приводит к разрыву пептидных связей и образованию ДНФ-производного Nконцевой аминокислоты Последнее экстрагируется эфиром и идентифицируется методом ТСХ в присутствии стандартов
 Дансильный метод l Разработан в 1963 г. английскими биохимиками В. Греем и Б. Хартли l Также основан на введении в аминогруппы белка «метки» , не удаляющейся при последующем гидролизе Брайен Хартли
Дансильный метод l Разработан в 1963 г. английскими биохимиками В. Греем и Б. Хартли l Также основан на введении в аминогруппы белка «метки» , не удаляющейся при последующем гидролизе Брайен Хартли
 Дансильный метод l Первая стадия – реакция дансилхлорида (1 -диметиламинонафталин-5 -сульфохлорида) с непротонированной -аминогруппой пептида (или белка) с образованием дансилпептида (ДНС-пептида)
Дансильный метод l Первая стадия – реакция дансилхлорида (1 -диметиламинонафталин-5 -сульфохлорида) с непротонированной -аминогруппой пептида (или белка) с образованием дансилпептида (ДНС-пептида)

 Дансильный метод l Полученный ДНС-белок подвергается гидролизу в присутствии 5, 7 н. НСl, при 105 С в течение 12– 16 ч, отщепившуюся ДНС-аминокислоту идентифицируют по флуоресценции в УФ-лучах
Дансильный метод l Полученный ДНС-белок подвергается гидролизу в присутствии 5, 7 н. НСl, при 105 С в течение 12– 16 ч, отщепившуюся ДНС-аминокислоту идентифицируют по флуоресценции в УФ-лучах


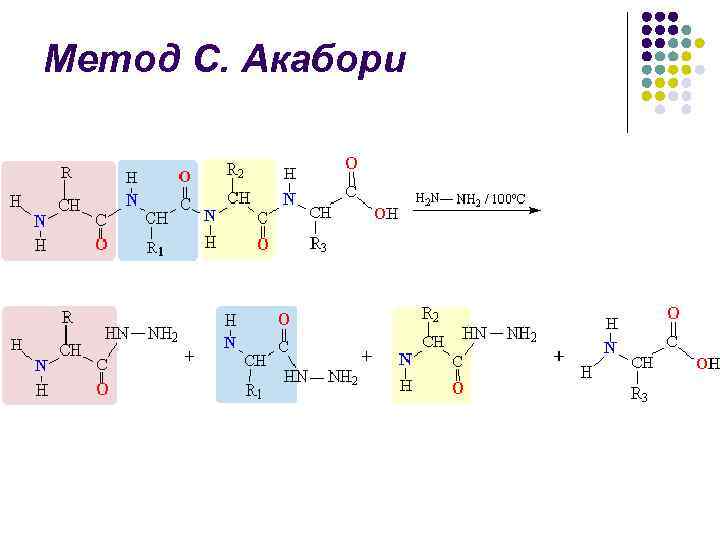 Метод С. Акабори l l При нагревании пептида (или белка) с безводным гидразином при 100– 120 С пептидные связи гидролизуются с образованием гидразидов аминокислот С-концевая аминокислота остается в виде свободной аминокислоты и может быть выделена из смеси и идентифицирована:
Метод С. Акабори l l При нагревании пептида (или белка) с безводным гидразином при 100– 120 С пептидные связи гидролизуются с образованием гидразидов аминокислот С-концевая аминокислота остается в виде свободной аминокислоты и может быть выделена из смеси и идентифицирована:

 Метод П. Эдмана l Метод деградации полипептидной цепи с помощью фенилизотиоционата (ФИТЦ) разработан шведским химиком П. В. Эдманом в 1950– 56 гг. l Метод Эдмана позволяет последовательно отщеплять N-концевые аминокислотные остатки в виде фенилтиогидантоинов (ФТГ) l Каждый цикл деградации включает 3 стадии
Метод П. Эдмана l Метод деградации полипептидной цепи с помощью фенилизотиоционата (ФИТЦ) разработан шведским химиком П. В. Эдманом в 1950– 56 гг. l Метод Эдмана позволяет последовательно отщеплять N-концевые аминокислотные остатки в виде фенилтиогидантоинов (ФТГ) l Каждый цикл деградации включает 3 стадии

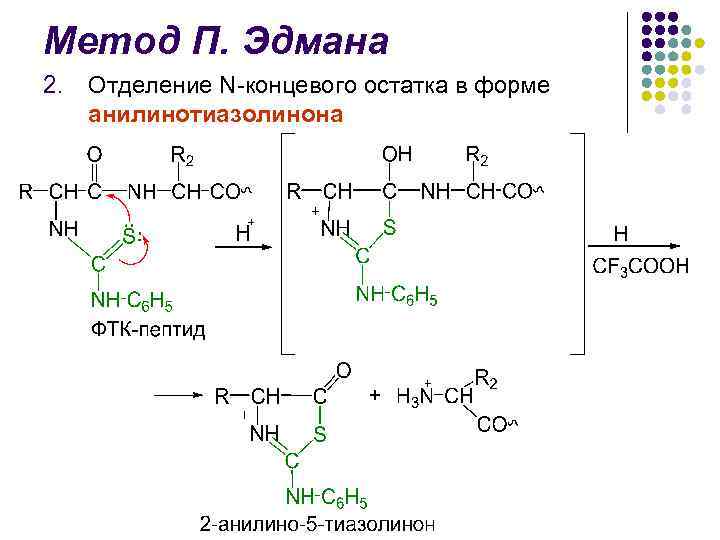
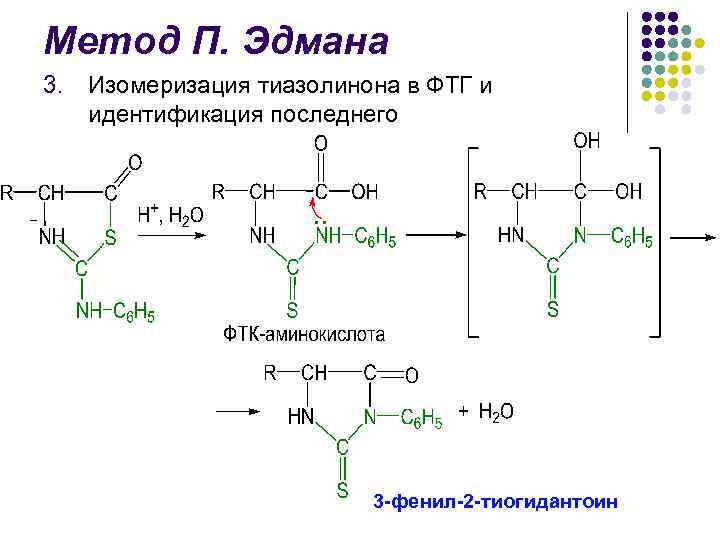

 Метод П. Эдмана l l l Идентификация отщепленных ФТГ является определяющей в методе Эдмана В течение длительного времени использовали хроматографию на бумаге, затем микротонкослойную хроматографию на силикагеле и полиамиде, жидкостную и газо-жидкостную хроматографии Крупным достижением в области структурных исследований белков явилось создание в 1967 г. П. Эдманом и Дж. Бэггом секвенатора (от англ. sequence – последовательность) – прибора, который с высокой эффективностью осуществляет последовательное автоматическое отщепление N-концевых аминокислотных остатков по методу Эдмана
Метод П. Эдмана l l l Идентификация отщепленных ФТГ является определяющей в методе Эдмана В течение длительного времени использовали хроматографию на бумаге, затем микротонкослойную хроматографию на силикагеле и полиамиде, жидкостную и газо-жидкостную хроматографии Крупным достижением в области структурных исследований белков явилось создание в 1967 г. П. Эдманом и Дж. Бэггом секвенатора (от англ. sequence – последовательность) – прибора, который с высокой эффективностью осуществляет последовательное автоматическое отщепление N-концевых аминокислотных остатков по методу Эдмана
 l Для определения структуры пептидов и белков можно применять: l l ферменты, катализирующие отщепление N- и С-концевых аминокислотных остатков – аминои карбоксипептидазы физико-химические методы, в частности, масс-спектрометрию
l Для определения структуры пептидов и белков можно применять: l l ферменты, катализирующие отщепление N- и С-концевых аминокислотных остатков – аминои карбоксипептидазы физико-химические методы, в частности, масс-спектрометрию
 Анализ данных по первичной структуре позволяет сделать следующие общие выводы l l Стабильность первичной структуры обеспечивается в основном пептидными связями; возможно участие и небольшого числа дисульфидных связей В полипептидной цепи могут быть обнаружены разнообразные комбинации аминокислот В полипептидах были открыты все мыслимые дипептиды В полипептидах относительно редки повторяющиеся последовательности
Анализ данных по первичной структуре позволяет сделать следующие общие выводы l l Стабильность первичной структуры обеспечивается в основном пептидными связями; возможно участие и небольшого числа дисульфидных связей В полипептидной цепи могут быть обнаружены разнообразные комбинации аминокислот В полипептидах были открыты все мыслимые дипептиды В полипептидах относительно редки повторяющиеся последовательности
 Анализ данных по первичной структуре позволяет сделать следующие общие выводы l l Каждый индивидуальный гомогенный белок характеризуется уникальной первичной структурой; часто замены аминокислот приводят не только к структурным перестройкам, но и к изменениям физикохимических свойств и биологических функций В некоторых белках (особенно ферментах), обладающих близкими свойствами, встречаются идентичные пептидные структуры (последовательности аминокислот), в особенности в областях их активных центров. Этот принцип структурного подобия наиболее типичен для ряда протеолитических ферментов – трипсина, химотрипсина и др.
Анализ данных по первичной структуре позволяет сделать следующие общие выводы l l Каждый индивидуальный гомогенный белок характеризуется уникальной первичной структурой; часто замены аминокислот приводят не только к структурным перестройкам, но и к изменениям физикохимических свойств и биологических функций В некоторых белках (особенно ферментах), обладающих близкими свойствами, встречаются идентичные пептидные структуры (последовательности аминокислот), в особенности в областях их активных центров. Этот принцип структурного подобия наиболее типичен для ряда протеолитических ферментов – трипсина, химотрипсина и др.

 Водородные связи В неполярном окружении энергия водородной связи –СО НN– составляет около 16, 7 к. Дж/моль, а повышение полярности среды снижает эту энергию
Водородные связи В неполярном окружении энергия водородной связи –СО НN– составляет около 16, 7 к. Дж/моль, а повышение полярности среды снижает эту энергию

 Гидрофобные взаимодействия l l Имеют энтропийную природу Связаны с тем, что неполярные заместители выталкиваются из воды и стремятся ограничить свой контакт с ней Напротив, вода стремится восстановить свое структурированное состояние и как бы принудительно группирует заместители в кластеры, обладающие минимумом энергии Вступают в основном неполярные боковые группы аминокислотных остатков
Гидрофобные взаимодействия l l Имеют энтропийную природу Связаны с тем, что неполярные заместители выталкиваются из воды и стремятся ограничить свой контакт с ней Напротив, вода стремится восстановить свое структурированное состояние и как бы принудительно группирует заместители в кластеры, обладающие минимумом энергии Вступают в основном неполярные боковые группы аминокислотных остатков

 Ван-дер-ваальсовы взаимодействия l l Складываются из дисперсионных сил притяжения атомов и сил взаимного отталкивания их электронных оболочек Энергетический вклад каждого контакта невелик (
Ван-дер-ваальсовы взаимодействия l l Складываются из дисперсионных сил притяжения атомов и сил взаимного отталкивания их электронных оболочек Энергетический вклад каждого контакта невелик (
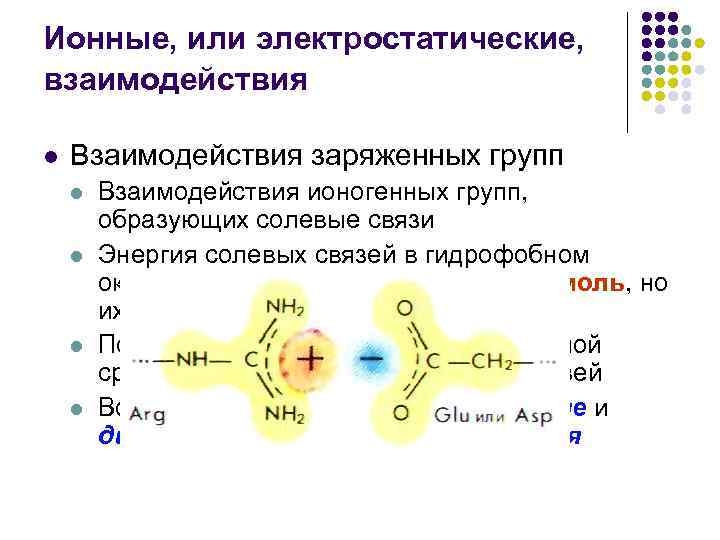 Ионные, или электростатические, взаимодействия l Взаимодействия заряженных групп l l Взаимодействия ионогенных групп, образующих солевые связи Энергия солевых связей в гидрофобном окружении может достигать 41, 9 к. Дж/моль, но их число сравнительно невелико Повышение диэлектрической постоянной среды понижает энергию солевых связей Во многом аналогичны ион-дипольные и диполь-дипольные взаимодействия
Ионные, или электростатические, взаимодействия l Взаимодействия заряженных групп l l Взаимодействия ионогенных групп, образующих солевые связи Энергия солевых связей в гидрофобном окружении может достигать 41, 9 к. Дж/моль, но их число сравнительно невелико Повышение диэлектрической постоянной среды понижает энергию солевых связей Во многом аналогичны ион-дипольные и диполь-дипольные взаимодействия
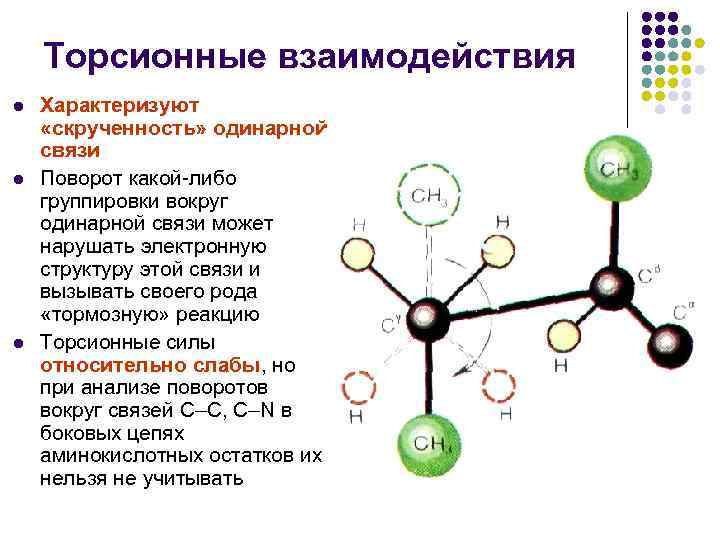 Торсионные взаимодействия l l l Характеризуют «скрученность» одинарной связи Поворот какой-либо группировки вокруг одинарной связи может нарушать электронную структуру этой связи и вызывать своего рода «тормозную» реакцию Торсионные силы относительно слабы, но при анализе поворотов вокруг связей С–С, С–N в боковых цепях аминокислотных остатков их нельзя не учитывать
Торсионные взаимодействия l l l Характеризуют «скрученность» одинарной связи Поворот какой-либо группировки вокруг одинарной связи может нарушать электронную структуру этой связи и вызывать своего рода «тормозную» реакцию Торсионные силы относительно слабы, но при анализе поворотов вокруг связей С–С, С–N в боковых цепях аминокислотных остатков их нельзя не учитывать

 Вторичная структура белка… l l пространственное расположение отдельных участков полипептидной цепи без учета типа и конформации боковых радикалов аминокислот Образуется за счет водородных связей между пептидными группами как одной цепи, так и разных цепей
Вторичная структура белка… l l пространственное расположение отдельных участков полипептидной цепи без учета типа и конформации боковых радикалов аминокислот Образуется за счет водородных связей между пептидными группами как одной цепи, так и разных цепей
 Вторичная структура белка l l l Любой участок молекулы белка имеет вторичную структуру Иногда рассматривают как вторичную структуру только периодические ее элементы: -спираль и -структуру Однако в белках встречаются участки, которые уложены вполне определенным способом, хотя их пространственная структура не содержит какого-либо периодически повторяющегося, регулярного мотива. Тем не менее к ним вполне приложимо понятие вторичной структуры Различают 2 вида вторичных структур: регулярные и нерегулярные Понятие вторичной структуры относится не ко всей белковой молекуле в целом, а к отдельным более или менее протяженным участкам ее полипептидной цепи
Вторичная структура белка l l l Любой участок молекулы белка имеет вторичную структуру Иногда рассматривают как вторичную структуру только периодические ее элементы: -спираль и -структуру Однако в белках встречаются участки, которые уложены вполне определенным способом, хотя их пространственная структура не содержит какого-либо периодически повторяющегося, регулярного мотива. Тем не менее к ним вполне приложимо понятие вторичной структуры Различают 2 вида вторичных структур: регулярные и нерегулярные Понятие вторичной структуры относится не ко всей белковой молекуле в целом, а к отдельным более или менее протяженным участкам ее полипептидной цепи
 Вторичная структура белка l Взаимодействия, которым принадлежит важнейшая роль в формировании вторичной структуры, определяются: l l l особенностями пептидной связи стерическими ограничениями (- и -углы) Основную роль играют водородные связи между периодически повторяющимися в цепи пептидными группировками
Вторичная структура белка l Взаимодействия, которым принадлежит важнейшая роль в формировании вторичной структуры, определяются: l l l особенностями пептидной связи стерическими ограничениями (- и -углы) Основную роль играют водородные связи между периодически повторяющимися в цепи пептидными группировками
 Стабилизация вторичной структуры за счет водородной связи l l Влияние окружающей белок воды Вода способна эффективно конкурировать за образование водородных связей: Влияние воды снижается при формировании компактной пространственной структуры белка, росте содержания пептидных связей, повышении вероятности их взаимодействия Таким образом, стабильность вторичной структуры зависит от ее включения в компактную третичную структуру
Стабилизация вторичной структуры за счет водородной связи l l Влияние окружающей белок воды Вода способна эффективно конкурировать за образование водородных связей: Влияние воды снижается при формировании компактной пространственной структуры белка, росте содержания пептидных связей, повышении вероятности их взаимодействия Таким образом, стабильность вторичной структуры зависит от ее включения в компактную третичную структуру

 -Спираль l В 50 -х годах ХХ в. Л. Полинг и Р. Кори, основываясь на данных о структуре кристаллов аминокислот и простых пептидов, рассмотрели возможные периодические конформации полипептидной цепи и пришли к выводу, что наиболее вероятна структура, названная ими -спиралью
-Спираль l В 50 -х годах ХХ в. Л. Полинг и Р. Кори, основываясь на данных о структуре кристаллов аминокислот и простых пептидов, рассмотрели возможные периодические конформации полипептидной цепи и пришли к выводу, что наиболее вероятна структура, названная ими -спиралью
 -Спираль В основу ее выбора были положены следующие критерии: 1. Образование плотноупакованной компактной структуры без пустот и перекрывания атомов 2. Максимальная насыщенность структуры водородными связями с тем условием, чтобы их геометрия была близка к линейной 3. Соблюдение межатомных расстояний и углов, свойственных аминокислотам и простым пептидам l
-Спираль В основу ее выбора были положены следующие критерии: 1. Образование плотноупакованной компактной структуры без пустот и перекрывания атомов 2. Максимальная насыщенность структуры водородными связями с тем условием, чтобы их геометрия была близка к линейной 3. Соблюдение межатомных расстояний и углов, свойственных аминокислотам и простым пептидам l
 -Спираль l С соблюдением этих условий можно построить как правую, так и левую спирали, однако правая -спираль оказывается энергетически несколько выгоднее левой, если пептидная цепь образована L-аминокислотами
-Спираль l С соблюдением этих условий можно построить как правую, так и левую спирали, однако правая -спираль оказывается энергетически несколько выгоднее левой, если пептидная цепь образована L-аминокислотами
 -Спираль l l Спиральная структура возникает, когда во всех звеньях полипептидной цепи углы поворота вокруг простых связей (и) имеют одинаковую величину и знак (близки к 60), что и приводит к постепенному закручиванию цепи При этом радикалы аминокислотных остатков оказываются на периферии образованного спиралью цилиндра и могут, в зависимости от характера их, обеспечивать гидрофобную или гидрофильную природу цилиндрической поверхности
-Спираль l l Спиральная структура возникает, когда во всех звеньях полипептидной цепи углы поворота вокруг простых связей (и) имеют одинаковую величину и знак (близки к 60), что и приводит к постепенному закручиванию цепи При этом радикалы аминокислотных остатков оказываются на периферии образованного спиралью цилиндра и могут, в зависимости от характера их, обеспечивать гидрофобную или гидрофильную природу цилиндрической поверхности

 -Спираль геометрические параметры: l l l l радиус r = 2, 3 Å (0, 23 нм) высота спирали (смещение) на 1 остаток d = 1, 5 Å шаг спирали (период идентичности) Р = 5, 4 Å 1 виток спирали образуют 3, 6 аминокислотных остатка все связи –С=О ориентированы вперед, к С-концу, а группы –N–Н – назад В -спирали каждая –NНгруппа соединена водородной связью с группой –СО четвертого от нее аминокислотного остатка (5 1 связь)
-Спираль геометрические параметры: l l l l радиус r = 2, 3 Å (0, 23 нм) высота спирали (смещение) на 1 остаток d = 1, 5 Å шаг спирали (период идентичности) Р = 5, 4 Å 1 виток спирали образуют 3, 6 аминокислотных остатка все связи –С=О ориентированы вперед, к С-концу, а группы –N–Н – назад В -спирали каждая –NНгруппа соединена водородной связью с группой –СО четвертого от нее аминокислотного остатка (5 1 связь)
 -Спираль l l Длина -спиральных участков в глобулярных белках относительно невелика (5– 15 аминокислотных остатков, 3 – 4 витка спирали), в фибриллярных белках – гораздо протяженнее Иногда наблюдаются изломы -спирали, обычно в местах включения остатков про, прерывающих системы водородных связей. При этом ось спирали отклоняется на 20– 30
-Спираль l l Длина -спиральных участков в глобулярных белках относительно невелика (5– 15 аминокислотных остатков, 3 – 4 витка спирали), в фибриллярных белках – гораздо протяженнее Иногда наблюдаются изломы -спирали, обычно в местах включения остатков про, прерывающих системы водородных связей. При этом ось спирали отклоняется на 20– 30

 -Структура l l Предложена У. Т. Астбери в 1941 г. на основании рентгеноструктурных исследований -кератина Через 10 лет Л. Полинг и Р. Кори установили, что структура, или «складчатый лист» , – это стабилизированный межцепочечными водородными связями ассоциат вытянутых, зигзагообразных пептидных цепей
-Структура l l Предложена У. Т. Астбери в 1941 г. на основании рентгеноструктурных исследований -кератина Через 10 лет Л. Полинг и Р. Кори установили, что структура, или «складчатый лист» , – это стабилизированный межцепочечными водородными связями ассоциат вытянутых, зигзагообразных пептидных цепей




 -Структура l l Число аминокислотных остатков в отрезке пептидной цепи, образующем -структуру, обычно от 3 до 8 Протяженная -структура, так называемый -слой, или -складчатый лист чаще всего состоит из 2– 6 цепей, иногда до 10
-Структура l l Число аминокислотных остатков в отрезке пептидной цепи, образующем -структуру, обычно от 3 до 8 Протяженная -структура, так называемый -слой, или -складчатый лист чаще всего состоит из 2– 6 цепей, иногда до 10
 -Структура l l l Боковые группы аминокислотных остатков при образовании структуры оказываются по разные стороны ее поверхности Сама же поверхность имеет складчатую форму, причем складки заданы -углеродными атомами Отходящие от них боковые группы образуют гребни Это позволяет формировать довольно протяженные поверхности, насыщенные однотипными (например, гидрофобными) боковыми радикалами Гидрофобные поверхности -складчатого слоя, взаимодействуя между собой или с гидрофобными гребнями -спиралей, участвуют в построении внутримолекулярных гидрофобных ядер, стабилизирующих пространственную структуру белка
-Структура l l l Боковые группы аминокислотных остатков при образовании структуры оказываются по разные стороны ее поверхности Сама же поверхность имеет складчатую форму, причем складки заданы -углеродными атомами Отходящие от них боковые группы образуют гребни Это позволяет формировать довольно протяженные поверхности, насыщенные однотипными (например, гидрофобными) боковыми радикалами Гидрофобные поверхности -складчатого слоя, взаимодействуя между собой или с гидрофобными гребнями -спиралей, участвуют в построении внутримолекулярных гидрофобных ядер, стабилизирующих пространственную структуру белка

 -Изгиб l l Как -спираль, так и -структура обычно представлены в глобулярных белках сравнительно короткими отрезками, поэтому значительная часть вторичной структуры белка приходится на разного рода петли, позволяющие изменить направление пептидной цепи Наиболее экономный структурный элемент, позволяющий повернуть полипептид на 180 , используя всего 3 пептидные группировки, – -изгиб Стабилизируется одной водородной связью Практически всегда оказывается на поверхности белковой глобулы, поэтому нередко играет существенную роль в ее взаимодействии с другими молекулами
-Изгиб l l Как -спираль, так и -структура обычно представлены в глобулярных белках сравнительно короткими отрезками, поэтому значительная часть вторичной структуры белка приходится на разного рода петли, позволяющие изменить направление пептидной цепи Наиболее экономный структурный элемент, позволяющий повернуть полипептид на 180 , используя всего 3 пептидные группировки, – -изгиб Стабилизируется одной водородной связью Практически всегда оказывается на поверхности белковой глобулы, поэтому нередко играет существенную роль в ее взаимодействии с другими молекулами


 l По результатам рентгеноструктурного анализа обнаружено, что способ укладки молекулы белка определяется его аминокислотной последовательностью l l l образованию -спирали способствуют ала, лей, глу образованию -структуры – мет, вал, иле изгибу цепи – гли, про, асн Если из 6 сгруппированных остатков аминокислот 4 способствуют образованию спирали, то данная группа – центр спирализации Если 3 остатка из 5 сгруппированных способствуют образованию -структуры, то данная группировка – затравка для -слоя
l По результатам рентгеноструктурного анализа обнаружено, что способ укладки молекулы белка определяется его аминокислотной последовательностью l l l образованию -спирали способствуют ала, лей, глу образованию -структуры – мет, вал, иле изгибу цепи – гли, про, асн Если из 6 сгруппированных остатков аминокислот 4 способствуют образованию спирали, то данная группа – центр спирализации Если 3 остатка из 5 сгруппированных способствуют образованию -структуры, то данная группировка – затравка для -слоя

 Сверхвторичная (надвторичная) структура белков l l Этот уровень организации белковой молекулы связан с наличием ансамблей взаимодействующих между собой вторичных структур Пространственное строение таких ансамблей и называют сверхвторичной структурой белковой молекулы, которая энергетически наиболее предпочтительна
Сверхвторичная (надвторичная) структура белков l l Этот уровень организации белковой молекулы связан с наличием ансамблей взаимодействующих между собой вторичных структур Пространственное строение таких ансамблей и называют сверхвторичной структурой белковой молекулы, которая энергетически наиболее предпочтительна

 Суперспирализованная -спираль l l Образуется при скручивании нескольких -спиралей в протофибриллы (-кератин), которые объединяются в микрофибриллы -Спирали удерживаются силами Ван-дер-Ваальса
Суперспирализованная -спираль l l Образуется при скручивании нескольких -спиралей в протофибриллы (-кератин), которые объединяются в микрофибриллы -Спирали удерживаются силами Ван-дер-Ваальса

 х l звено, состоящее из двух параллельных -слоев с сочленением между ними в виде: l неупорядоченного клубка – с
х l звено, состоящее из двух параллельных -слоев с сочленением между ними в виде: l неупорядоченного клубка – с
 х l звено, состоящее из двух параллельных -слоев с сочленением между ними в виде: l l -спирали – – укладка цепи по Россману Два последовательно соединенных участка образуют структуру «греческий орнамент» –
х l звено, состоящее из двух параллельных -слоев с сочленением между ними в виде: l l -спирали – – укладка цепи по Россману Два последовательно соединенных участка образуют структуру «греческий орнамент» –
 х l звено, состоящее из двух параллельных -слоев с сочленением между ними в виде: l -структуры – – -зигзаг, «греческий ключ»
х l звено, состоящее из двух параллельных -слоев с сочленением между ними в виде: l -структуры – – -зигзаг, «греческий ключ»
 Домены l l Многие белки содержат относительно слабо взаимодействующие между собой участки – домены Домены (от фр. domaine – владение) – структурно и функционально обособленные области (субобласти) молекулы, соединенные друг с другом короткими участками полипептидной цепи, которые называются шарнирными участками Функциональные домены могут состоять из одного или нескольких структурных доменов У ряда ферментов в углублениях между доменами располагается активный центр
Домены l l Многие белки содержат относительно слабо взаимодействующие между собой участки – домены Домены (от фр. domaine – владение) – структурно и функционально обособленные области (субобласти) молекулы, соединенные друг с другом короткими участками полипептидной цепи, которые называются шарнирными участками Функциональные домены могут состоять из одного или нескольких структурных доменов У ряда ферментов в углублениях между доменами располагается активный центр


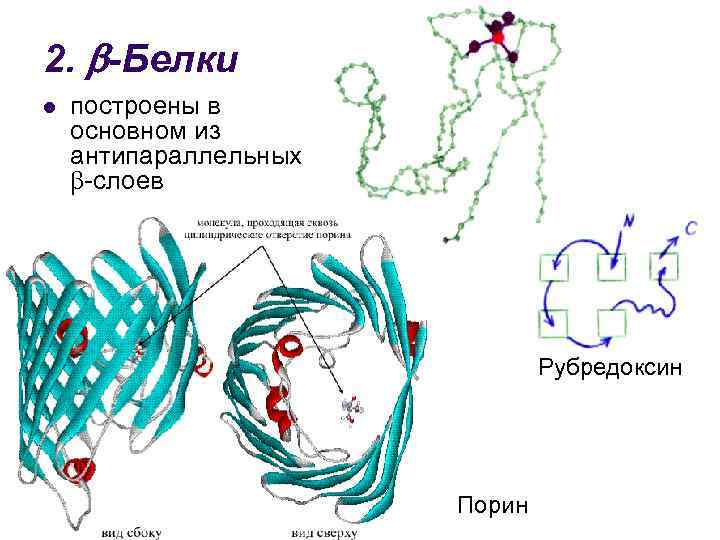
 3. + -Белки l имеются участки, целиком построенные из -спиралей, и участки, целиком построенные из антипараллельных -слоев Инсулин
3. + -Белки l имеются участки, целиком построенные из -спиралей, и участки, целиком построенные из антипараллельных -слоев Инсулин

 4. / -Белки l l -Спирали и -структуры чередуются по ходу цепи Большинство -структур (параллельные) оказываются локализованными в центральной части молекулы, где эти структуры изгибаются в виде пропеллера («твист» структуры), образуя жесткую «основу» , с которой связаны остальные участки молекулы
4. / -Белки l l -Спирали и -структуры чередуются по ходу цепи Большинство -структур (параллельные) оказываются локализованными в центральной части молекулы, где эти структуры изгибаются в виде пропеллера («твист» структуры), образуя жесткую «основу» , с которой связаны остальные участки молекулы


 Домены l l l Средний размер домена обычно – 100– 150 остатков, что отвечает глобуле с поперечником около 2, 5 нм Вместе с тем встречаются и значительно большие домены Вероятнее всего, формирование пространственной структуры белка вначале происходит внутри будущих доменов, а взаимная укладка доменов, т. е. образование третичной структуры, происходит на заключительных этапах формирования глобулы
Домены l l l Средний размер домена обычно – 100– 150 остатков, что отвечает глобуле с поперечником около 2, 5 нм Вместе с тем встречаются и значительно большие домены Вероятнее всего, формирование пространственной структуры белка вначале происходит внутри будущих доменов, а взаимная укладка доменов, т. е. образование третичной структуры, происходит на заключительных этапах формирования глобулы

 Третичная структура белка… l l l характеризует пространственное расположение упорядоченных и аморфных участков в полипептидной цепи в целом, которое достигается за счет взаимодействия боковых радикалов и зависит от их типа и конформации Таким образом, третичная структура описывает пространственную укладку всей молекулы белка, если она образована одной полипептидной цепью Имеет прямое отношение к форме молекул белка, которая может быть различной: от шарообразной до нитевидной
Третичная структура белка… l l l характеризует пространственное расположение упорядоченных и аморфных участков в полипептидной цепи в целом, которое достигается за счет взаимодействия боковых радикалов и зависит от их типа и конформации Таким образом, третичная структура описывает пространственную укладку всей молекулы белка, если она образована одной полипептидной цепью Имеет прямое отношение к форме молекул белка, которая может быть различной: от шарообразной до нитевидной
 Нитевидные, или фибриллярные белки l l фиброин шелка кератин волос, рогов, копыт коллаген и др.
Нитевидные, или фибриллярные белки l l фиброин шелка кератин волос, рогов, копыт коллаген и др.

 Третичная структура белка l l основа функциональности белка, которая требует точной пространственной организации больших ансамблей, построенных из множества аминокислотных остатков Такие ансамбли (домены) формируют: l l активные центры ферментов зоны связывания других биологических молекул эффекторные центры белков и т. д. , поэтому нарушение третичной структуры белка (денатурация) неизменно приводит к утрате им способности функционировать
Третичная структура белка l l основа функциональности белка, которая требует точной пространственной организации больших ансамблей, построенных из множества аминокислотных остатков Такие ансамбли (домены) формируют: l l активные центры ферментов зоны связывания других биологических молекул эффекторные центры белков и т. д. , поэтому нарушение третичной структуры белка (денатурация) неизменно приводит к утрате им способности функционировать
 Третичная структура белка l l l Стабильность третичной структуры зависит от системы нековалентных взаимодействий внутри белковой глобулы (?) Некоторые белки дополнительно стабилизируются ковалентными – дисульфидными – связями Gцепь = RТln. К = Нцепь – Т Sцепь Нцепь 0 (S
Третичная структура белка l l l Стабильность третичной структуры зависит от системы нековалентных взаимодействий внутри белковой глобулы (?) Некоторые белки дополнительно стабилизируются ковалентными – дисульфидными – связями Gцепь = RТln. К = Нцепь – Т Sцепь Нцепь 0 (S
 Третичная структура белка l Соотношение Нцепь и –Т Sцепь зависит от: l l числа нековалентных связей, устанавливающихся при свертывании молекулы белка в глобулу, а значит, от длины полипептидной цепи от температуры. С ростом Т |Т S| рано или поздно превысит | Н| и пространственная структура утратит стабильность – тепловая денатурация белка
Третичная структура белка l Соотношение Нцепь и –Т Sцепь зависит от: l l числа нековалентных связей, устанавливающихся при свертывании молекулы белка в глобулу, а значит, от длины полипептидной цепи от температуры. С ростом Т |Т S| рано или поздно превысит | Н| и пространственная структура утратит стабильность – тепловая денатурация белка
 Третичная структура белка l l При свертывании глобулы убывает энтропия пептидной цепи, но одновременно происходит возрастание энтропии растворителя – воды Это играет решающую роль в стабилизации третичной структуры
Третичная структура белка l l При свертывании глобулы убывает энтропия пептидной цепи, но одновременно происходит возрастание энтропии растворителя – воды Это играет решающую роль в стабилизации третичной структуры
 Третичная структура белка l l Вода – необычный, в высокой мере структурированный растворитель (?) Полярные группы белка легко включаются в льдоподобные структуры воды, устанавливая с ней водородные связи Неполярные группы занимают своеобразные полости, образованные молекулами воды, связанными водородными связями Все это выключает из хаотического движения значительное число молекул воды, т. е. происходит упорядочение, уменьшение энтропии воды
Третичная структура белка l l Вода – необычный, в высокой мере структурированный растворитель (?) Полярные группы белка легко включаются в льдоподобные структуры воды, устанавливая с ней водородные связи Неполярные группы занимают своеобразные полости, образованные молекулами воды, связанными водородными связями Все это выключает из хаотического движения значительное число молекул воды, т. е. происходит упорядочение, уменьшение энтропии воды
 Третичная структура белка l l l Но так как неполярные группы белка не могут «отслаиваться» от молекулы, происходит свертывание глобулы, при котором значительная часть (не менее ½) гидрофобных групп оказывается скрытой от контакта с водой Это влечет установление гидрофобных контактов, ван-дер-ваальсовых сил Происходит понижение энтропии белка Следствием образования глобулы является разрушение значительной части водородных связей и гидрофобных взаимодействий боковых групп с молекулами воды Возрастает хаотичность движения последних, и энтропия воды растет
Третичная структура белка l l l Но так как неполярные группы белка не могут «отслаиваться» от молекулы, происходит свертывание глобулы, при котором значительная часть (не менее ½) гидрофобных групп оказывается скрытой от контакта с водой Это влечет установление гидрофобных контактов, ван-дер-ваальсовых сил Происходит понижение энтропии белка Следствием образования глобулы является разрушение значительной части водородных связей и гидрофобных взаимодействий боковых групп с молекулами воды Возрастает хаотичность движения последних, и энтропия воды растет

 Четвертичная структура белка… l l l Четвертичную структуру имеют олигомерные белки, молекулы которых состоят из двух и более полипептидных цепей, связанных нековалентно Четвертичная структура – размещение в пространстве взаимодействующих между собой субъединиц, образованных отдельными полипептидными цепями белка Взаимодействие между субъединицами достаточно сильно, так что их ансамбль (ансамбль глобул) выступает как единая молекула, в то же время каждая из объединившихся глобул сохраняет значительную автономность
Четвертичная структура белка… l l l Четвертичную структуру имеют олигомерные белки, молекулы которых состоят из двух и более полипептидных цепей, связанных нековалентно Четвертичная структура – размещение в пространстве взаимодействующих между собой субъединиц, образованных отдельными полипептидными цепями белка Взаимодействие между субъединицами достаточно сильно, так что их ансамбль (ансамбль глобул) выступает как единая молекула, в то же время каждая из объединившихся глобул сохраняет значительную автономность

 Четвертичная структура белка l l Субъединицы, образующие четвертичную структуру белка, могут быть совершенно различными по строению и функциям – гетеромерные белки Это позволяет объединить в одной структуре несколько взаимосвязанных функций, создать полифункциональную молекулу l l Пример. Протеинкиназа: С-субъединица отвечает за ферментативную активность, а R-субъединица – регуляторная В гомомерных белках субъединицы одинаковы
Четвертичная структура белка l l Субъединицы, образующие четвертичную структуру белка, могут быть совершенно различными по строению и функциям – гетеромерные белки Это позволяет объединить в одной структуре несколько взаимосвязанных функций, создать полифункциональную молекулу l l Пример. Протеинкиназа: С-субъединица отвечает за ферментативную активность, а R-субъединица – регуляторная В гомомерных белках субъединицы одинаковы
 Четвертичная структура белка l Межсубъединичные контакты – весьма развитая система нековалентных взаимодействий l l Контакты между гидрофобными группами и целыми гидрофобными участками на поверхности субъединиц (контактные площадки, или зоны), приводящие к возрастанию энтропии системы и сдвигу равновесия в сторону олигомера водородные связи электростатические взаимодействия между боковыми группами Четвертичная структура менее прочная, чем третичная, т. к. меньше вклад гидрофобных контактов
Четвертичная структура белка l Межсубъединичные контакты – весьма развитая система нековалентных взаимодействий l l Контакты между гидрофобными группами и целыми гидрофобными участками на поверхности субъединиц (контактные площадки, или зоны), приводящие к возрастанию энтропии системы и сдвигу равновесия в сторону олигомера водородные связи электростатические взаимодействия между боковыми группами Четвертичная структура менее прочная, чем третичная, т. к. меньше вклад гидрофобных контактов

 Функции четвертичной структуры 1. Объединение нескольких взаимосвязанных функций в единой структуре 2. Архитектурная функция 1. Формирование пространственных образований весьма сложной конфигурации, обеспечивающих специфические функциональные возможности белка (ферритин) 2. Суммарное проведение последовательных реакций ферментами 3. Формирование функциональных центров (активных центров ферментов)
Функции четвертичной структуры 1. Объединение нескольких взаимосвязанных функций в единой структуре 2. Архитектурная функция 1. Формирование пространственных образований весьма сложной конфигурации, обеспечивающих специфические функциональные возможности белка (ферритин) 2. Суммарное проведение последовательных реакций ферментами 3. Формирование функциональных центров (активных центров ферментов)
 Функции четвертичной структуры 3. Обеспечение множественных взаимодействий белка с протяженными структурами l ДНК-связывающие белки – димеры (?) 4. Регуляторная функция. Передача эффекта (нарушение третичной структуры при взаимодействии с субстратом) передается от одной субъединицы к другой, что приводит к перестройке всей четвертичной структуры
Функции четвертичной структуры 3. Обеспечение множественных взаимодействий белка с протяженными структурами l ДНК-связывающие белки – димеры (?) 4. Регуляторная функция. Передача эффекта (нарушение третичной структуры при взаимодействии с субстратом) передается от одной субъединицы к другой, что приводит к перестройке всей четвертичной структуры


 1. Каждый индивидуальный белок характеризуется уникальной структурой, обеспечивающей уникальность его функций 2. Поэтому выяснение структуры разнообразных белков может служить ключом к познанию природы живых систем и, соответственно, сущности жизни
1. Каждый индивидуальный белок характеризуется уникальной структурой, обеспечивающей уникальность его функций 2. Поэтому выяснение структуры разнообразных белков может служить ключом к познанию природы живых систем и, соответственно, сущности жизни
 Литература l l l l Березов Т. Т. , Коровкин Б. Ф. Биологическая химия. – М. : Медицина, 1983 Мецлер Д. Биохимия. Химические реакции в живой клетке. В 3 -х т. – Т. т. 1, 2. – М. : Мир, 1980 Овчинников Ю. А. Биоорганическая химия. – М. : Просвещение, 1987 Основы биохимии / Под ред. А. А. Анисимова. – М. : Высшая школа, 1986 Рис Э. , Стренберг М. От клеток к атомам. Иллюстрированное введение в молекулярную биологию. – М. : Мир, 1988 Степанов В. М. Молекулярная биология. Структура и функции белков. – М. : Высшая школа, 1996 Филиппович Ю. Б. Основы биохимии. – М. : Высшая школа, 1993
Литература l l l l Березов Т. Т. , Коровкин Б. Ф. Биологическая химия. – М. : Медицина, 1983 Мецлер Д. Биохимия. Химические реакции в живой клетке. В 3 -х т. – Т. т. 1, 2. – М. : Мир, 1980 Овчинников Ю. А. Биоорганическая химия. – М. : Просвещение, 1987 Основы биохимии / Под ред. А. А. Анисимова. – М. : Высшая школа, 1986 Рис Э. , Стренберг М. От клеток к атомам. Иллюстрированное введение в молекулярную биологию. – М. : Мир, 1988 Степанов В. М. Молекулярная биология. Структура и функции белков. – М. : Высшая школа, 1996 Филиппович Ю. Б. Основы биохимии. – М. : Высшая школа, 1993
Биосинтез белка.
1. Структура одного белка определяется:
1)группой генов 2)одним геном
3)одной молекулой ДНК 4)совокупностью генов организма
2. Ген кодирует информацию о последовательности мономеров в молекуле:
1)т-РНК 2) АК 3) гликогена 4) ДНК
3. Антикодонами называются триплеты:
1)ДНК 2) т-РНК 3)и-РНК 4) р-РНК
4. Пластический обмен состоит преимущественно из реакций:
1)распада органических веществ 2)распада неорганических веществ
3)синтеза органических веществ 4)синтеза неорганических веществ
5. Синтез белка в прокариотической клетке происходит:
1)на рибосомах в ядре 2)на рибосомах в цитоплазме 3)в клеточной стенке
4)на внешней поверхности цитоплазматической мембраны
6. Процесс трансляции происходит:
1)в цитоплазме 2)в ядре 3)в митохондриях
4)на мембранах шероховатой эндоплазматической сети
7. На мембранах гранулярной эндоплазматической сети происходит синтез:
1)АТФ; 2)углеводов; 3) липидов; 4)белков.
8. Один триплет кодирует:
1.одну АК 2 один признак организма 3. несколько АК
9. Синтез белка завершается в момент
1.узнавание кодона антикодоном 2.появление на рибосоме «знака препинания»
3.поступление и-РНК на рибосому
10. Процесс, в результате которого происходит считывание информации с молекулы ДНК.
1.трансляция 2.транскрипция 3.трансформация
11. Свойства белков определяется…
1.вторичной структурой белка 2.первичной структурой белка
3.третичной структурой белка
12. Процесс, при котором антикодон узнаёт кодон на и-РНК
13. Этапы биосинтеза белка.
1.транскрипция, трансляция 2.трансформация, трансляция
3.трансорганизация, транскрипция
14. Антикодон т-РНК состоит из нуклеотидов УЦГ. Какой триплет ДНК ему комплементарен ?
1.УУГ 2. ТТЦ 3. ТЦГ
15. Количество т-РНК, участвующих в трансляции, равно количеству:
1.Кодонов и-РНК, шифрующих аминокислоты 2. Молекул и-РНК
3 Генов, входящих в молекулу ДНК 4. Белков, синтезируемых на рибосомах
16. Установите последовательность расположения нуклеотидов и-РНК при транскрипции с одной из цепей ДНК: А-Г-Т-Ц-Г
1) У 2) Г 3) Ц 4) А 5) Ц
17. При репликации молекулы ДНК образуется:
1) нить, распавшаяся на отдельные фрагменты дочерних молекул
2) молекула, состоящая из двух новых цепей ДНК
3) молекула, половина которой состоит из нити иРНК
4) дочерняя молекула, состоящая из одной старой и одной новой цепи ДНК
18. Матрицей для синтеза молекулы иРНК при транскрипции служит:
1) вся молекула ДНК 2) полностью одна из цепей молекулы ДНК
3) участок одной из цепей ДНК
4) в одних случаях одна из цепей молекулы ДНК, в других– вся молекула ДНК.
19.Процесс самоудвоения молекулы ДНК.
1.репликация 2.репарация
3. реанкорнация
20. При биосинтезе белка в клетке энергия АТФ:
1) расходуется 2) запасается
3) не расходуется и не выделяется
21. В соматических клетках многоклеточного организма:
1) различный набор генов и белков 2) одинаковый набор генов и белков
3) одинаковый набор генов, но разный набор белков
4) одинаковый набор белков, но разный набор генов
22.. Один триплет ДНК несет информацию о:
1)последовательности аминокислот в молекуле белка
2)признаке организма 3)аминокислоте в молекуле синтезируемого белка
4)составе молекулы РНК
23. Какой из процессов не происходит в клетках любого строения и функции:
1)синтез белков 2) обмен веществ 3)митоз 4) мейоз
24. Понятие «транскрипция» относится к процессу:
1)удвоения ДНК 2)синтеза и-РНК на ДНК
3)перехода и-РНК на рибосомы 4)создания белковых молекул на полисоме
25. Участок молекулы ДНК, несущий информацию об одной молекуле белка - это:
1)ген 2)фенотип 3)геном 4)генотип
26. Транскрипция у эукариот происходит в:
1)цитоплазме 2)эндоплазматической мембране 3)лизосомах 4)ядре
27.Синтез белка происходит в:
1)гранулярном эндоплазматическом ретикулуме
2)гладком эндоплазматическом ретикулуме 3)ядре 4) лизосомах
28. Одна аминокислота кодируется:
1)четырьмя нуклеотидами 2)двумя нуклеотидами
3)одним нуклеотидом 4) тремя нуклеотидами
29. Триплету нуклеотидов АТЦ в молекуле ДНК будет соответствовать кодон молекулы и-РНК:
1) ТАГ 2) УАГ 3) УТЦ 4) ЦАУ
30. Знаки препинания генетического кода:
1.кодируют определённые белки 2. запускают синтез белка
3. прекращают синтез белка
31. Процесс самоудвоения молекулы ДНК.
1.репликация 2.репарация 3.реанкорнация
32. Функция и-РНК в процессе биосинтеза.
1.хранение наследственной информации 2.транспорт АК на рибосомы
3.подача информации на рибосомы
33. Процесс, когда т-РНК приносят аминокислоты на рибосомы.
1.транскрипция 2.трансляция 3.трансформация
34. Рибосомы, синтезирующие одну и ту же белковую молекулу.
1.хромосома 2.полисома 3.мегахромосома
35. Процесс, при котором аминокислоты образуют белковую молекулу.
1.транскрипция 2.трансляция 3.трансформация
36. К реакциям матричного синтеза относят…
1.репликацию ДНК 2.транскрипцию, трансляцию 3.оба ответа правильные
37.Один триплет ДНК несет информацию о:
1.Последовательности аминокислот в молекуле белка
2.Месте определенной АК в белковой цепи
3.Признаке конкретного организма
4.Аминокислоте, включаемой в белковую цепь
38. В гене закодирована информация о:
1) строении белков, жиров и углеводов 2) первичной структуре белка
3) последовательности нуклеотидов в ДНК
4) последовательности аминокислот в 2-х и более молекулах белков
39. Синтез иРНК начинается с:
1) разъединения ДНК на две нити 2) взаимодействия фермента РНК - полимеразы и гена
3) удвоения гена 4) распада гена на нуклеотиды
40. Транскрипция происходит:
1) в ядре 2) на рибосомах 3) в цитоплазме 4) на каналах гладкой ЭПС
41. Синтез белка не идет на рибосомах у:
1) возбудителя туберкулеза 2) пчелы 3) мухомора 4) бактериофага
42. При трансляции матрицей для сборки полипептидной цепи белка служат:
1) обе цепочки ДНК 2) одна из цепей молекулы ДНК
3) молекула иРНК 4) в одних случаях одна из цепей ДНК, в других– молекула иРНК